Turmeric, Cinnamon and Cloves Inhibits Effect Of iNOS/COX2 And Nitrite Production in Inflamed Murine Macrophages
This was my bachelor’s thesis, marking my first exposure to scientific exploration, from designing to writing. 😍😍😍 Written in 2019. Type of writing: Research. Complexity: III
Dhruvini
3/4/2026
Abstract
Inflammation is one of the key defences against pathogen or tissue injury essential for human survival. It should be short-lived but sometimes due to pathological circumstances, the process doesn't terminate and eventually develops into chronic inflammation. iNOS/COX2 enzymes have been shown to upregulate in inflammatory diseases. iNOS is used in the production of NO while COX2 is used in the production of PGE2. Both have a various role such as dilation of blood vessel thus aiding in the recruitment of leukocytes. Upregulation of these molecules enhances inflammatory response that leads to tissue injury and tumour formation.
The main aim of the research is to find out if turmeric, cinnamon, and cloves have an inhibitory effect on iNOS/COX2 (using western blot) and decrease NO levels (measuring nitrite level via Griess assay) in macrophages (J774 cells). The compounds were essential oil dissolve in ethanol. Cell viability using trypan blue and resazurin assay was carried out to determine maximum dilution of compounds used for Griess assay and Western blot. Cell lysis (using SDS) was used for protein extraction and BCA assay was used to determine the volume of cell lysate to be loaded for western blot. Western blot was used to determine protein expression.
All compounds have expressed the inhibitory effect for iNOS/COX2 and decreased the nitrite levels compared to inflamed cells. Turmeric has shown to exhibit the most anti-inflammatory properties, with nitrite levels decreased by 27.2µM while cloves show the least, with nitrite levels 13.3µM. This may be due to its turmeric ability to impact the enzymes directly via enhancing its degradation by proteasome or ubiquitination. Cinnamon has the most inhibitory effect on iNOS (due to its ability to impact NF-kb proteins) but had a higher nitrite levels than expected. One explanation of its upregulation of eNOS and nNOS. Overall further research is required especially in a vivo and clinical trial to determine its ability to be used in alternative medicine against chronic inflammation.
Introduction
Inflammation in humans is the body defence mechanism initiated by the immune response to any foreign substances including infectious microorganism, an object or toxic compound that damages cells (Cavaillon & Singer,2011). Inflammation has a complex molecular pathway with variation depending on the stimuli but shares the same key processes. These involve activation of Pattern recognition receptor (PRR), activation of inflammatory pathway, the release of inflammatory mediators and engagement of inflammatory cells.
1.1. Activation of PRR
PRR is expressed by all somatic cells and consist of different classes including Toll-like receptors (TLRs), C-type lectin receptors (CLRs), retinoic acid-inducible gene-I-like receptors (RLRs), and NOD-like receptors (NLRs). In humans, TLRs are the largest family and the most researched class of PRR (L. Chen et al., 2017). PRR can recognize and bind to pathogen-associated molecular pattern (PAMPS) attach on the pathogen few examples are TLRs2 can recognize cell surface components such as glycoprotein on virus envelope, peptidoglycan on bacteria, GPI-mucin on protozoa, Zymosan and β-Glycan on fungi (Mogensen, 2009). TLRs4 can bind to danger-associated molecular patterns (DAMPS) which are biomolecules that are released during cell damage.
1.2. Activation of Signalling pathway
Activation of PRR leads to dimerization that trigger the 3 main inflammatory pathway that compromises of Janus protein-tyrosine kinase/signal transducer and activator of transcription (JAK-STAT), nuclear factor kappa B (NF-kb) and Mitogen-activated protein kinase (MAPK). The pathways can be moderated by myeloid differentiation factor-88 (MyD88) or TIR-domain-containing adapter-inducing interferon-β (TRIF)(Shu, Chen, He, Zhou, & Liang, 2016). Inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX2) are transcribed by NF-kb pathway and it is divided into 3 including canonical, non-canonical, and atypical IkB kinase (IKK) independent pathways summarized in Figure 1.
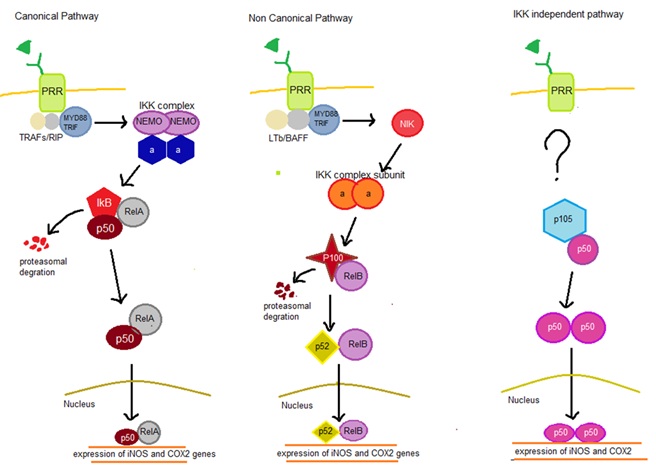
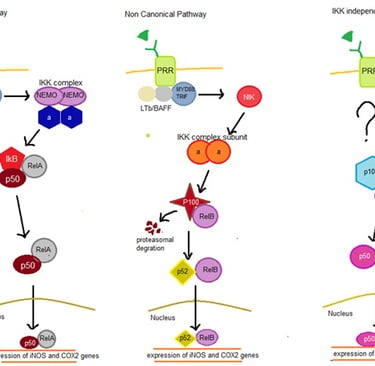
Figure 1: Graphical presentation of 3 different types of NF-KB Pathway
Normally. in canonical pathway p50/p65-Rel A complex is bound by inhibitor KB (IkB) inhibitory protein. Upon dimerization of PRR will trigger TNF Receptor-Associated Factor (TRAFs) and ribosome inactivating protein (RIP) to bind to the cytosolic domain of the PRR, these causes further binding of MyD88/TRIF resulting in further binding of IKK complex (consist of 2 NF-kappa-B essential modulators (NEMO) and 2 alpha subunit) resulting in degradation of IkB releasing p50/p65-Rel A complex which enter the nucleus to promote iNOS and COX-2 gene expression. In the non-conical pathway, binding of Lymphotoxin B (LTb) and B-cell activating factor (BAFF) to the cytosolic domain of PRR causes binding of MyD88/TRIF resulting in phosphorylation of NF-kB-inducing kinase (NIK) that triggers IKK subunit (consisting only alpha) leading to degradation of p100 in the P100/Rel B complex. These lead to the formation of p52/RelB complex that enters the nucleus for transcription. For the IKK independent pathway, the main signalling molecules is still unknown. During these process p105-p50/p52 undergoes processing forming a homodimer of p50 or p52 for transcription of iNOS and COX2 (Gilmore, 2006).
1.3. Inflammatory cytokines and recruitment of immune cells
Due to various signaling during inflammation many inflammatory molecules are released such as cytokines which play a key role in recruitment of leukocytes eg Interleukin-8 (IL-8), causes cytotoxic effect at the site of injury interferon-gamma (IFNγ), regulate inflammation and immune response eg tumor necrosis factor alpha (TNFα) and initiate haematopoiesis eg Interleukin-3 (IL-3) (Foster, 2001). Inflammatory proteins are also transcribed such as C-reactive protein (CRP) to activate the complement system (Du Clos, 2000) and haptoglobin to minimize iron loss and exert an antibacterial activity (Wassell, 2000).
1.4. The physiological function of iNOS
A major role of iNOS includes the production of Nitric oxide (NO) and these processes are calcium-independent unlike Endothelial nitric oxide synthase (eNOS) and Neuronal nitric oxide synthase nNOS. iNOS transfers electrons from Nicotinamide Adenine Dinucleotide Phosphate Hydrogen (NADPH) to oxygen molecule for the formation of free radicle oxygen species to oxidizes L-arginine to L-citrulline with oxygenase bound to cofactors such as flavin adenine dinucleotide (FAD), flavin mononucleotide (FMN), and (6R-)5,6,7,8-tetrahydrobiopterin (BH4). These leads to the synthesis of NO and (Nicotinamide adenine dinucleotide phosphate) NADP (Förstermann & Sessa, 2012). NO can freely move out from luminal sides of the endothelial cell into vascular smooth muscle and triggers guanylate cyclase causing the production of intracellular cyclic guanosine monophosphate (cGMP) resulting in relaxation of smooth vascular muscles leading to dilation of the lumen in a blood vessel. NO decreases the permeability of endothelium preventing diffusion of molecules such as histamine and platelet activating factor (PAF) between interstitium and the lumen of blood vessels. NO also inhibits the activity of b2 adhesion of neutrophils and downregulates p-selectin (binds to l-selectin on leukocytes casing leukocytes to roll down along the endothelial layer) expression on the endothelium. These both factors result in detachment of leukocytes to the endothelium increasing its velocity thus aiding in recruitment. NO decreases platelet activation and aggregation thus inhibiting the production of proinflammatory molecules such as serotonin, thromboxane, and lipoxins. NO can have an inhibitory effect on heme-containing enzymes such as mitochondrial electron transfer and cytochrome P450 preventing apoptosis in the cell (Wallace, 2005).
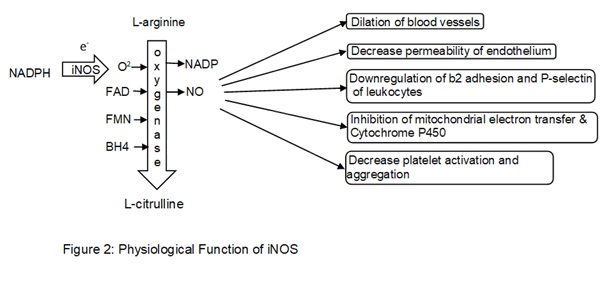
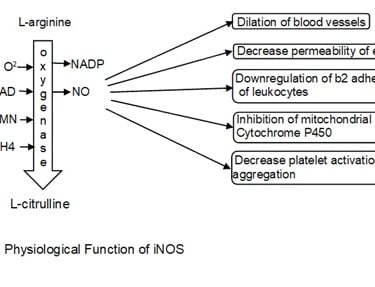
1.5. The physiological function of COX2
Arachidonic acid, released from the cell membrane by phospholipases (PLAs) are metabolized by COX2 resulting in production of Prostaglandin H2(PGH2) and by binding tissue-specific isomerase that act as a substrate produces molecules such as Prostaglandins and thromboxane A2 (TXA2), prostaglandin E2 (PGE2), prostacyclin (PGI2), prostaglandin D2 (PGD2) and prostaglandin F2α (PGF2α). PGE2 is the most abundant Prostaglandin produced in the body, and mediator of inflammation. It requires cofactors such as Cytosolic prostaglandin E2 synthase (cPGES) or Microsomal prostaglandin E synthase-1 (mPGES-1) and Microsomal prostaglandin E synthase-2 (mPGES-2) (Ricciotti & FitzGerald, 2011). PGE2 acts as a vasodilator and formation of edema, it also triggers sensory neuron initiating pain and initiate neuron in the preoptic area in the hypothalamus inducing fever. It also activates T Helper 17 Cell (TH17) which are known to produce interleukin-17 (IL-17) to promote the recruitment of leukocytes at the site of inflammation. PGE2 demises the expression of interleukin-2 (IL2) and IL2 receptors preventing proliferation and activation of T cells and promoting class switching from TH1 to TH2 response which exerts less tissue damage. PGE2 also downregulates the expression of Chemokine ligand 19 (CCL19) in monocytes and dendritic cells leading to inhibition of recruitment of naïve T cell and its progression to an effector T cell (Nakanishi & Rosenberg, 2013).
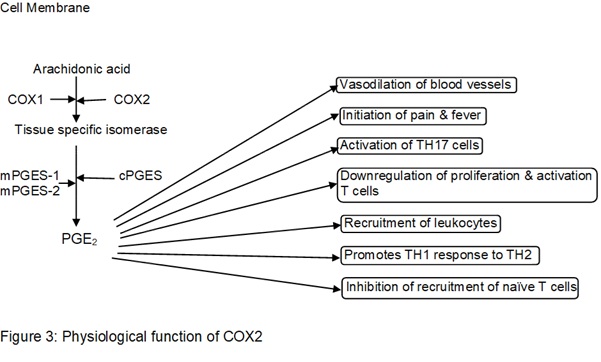
1.6. The consequence of unregulated inflammation
Inflammation should be short-lived and the process should dismiss once tissue repair has taken place but sometimes unregulated inflammation can lead to many chronic inflammatory diseases (defined as persistent inflammation occurs lasting from months to years) that lead to diseases such as diabetes, cardiovascular diseases, asthma, arthritis, neurodegenerative disorders such as Alzheimer's disease, etc (Straub & Schradin, 2016). According to WHO, during 2000 in united states 125 million individual are sufferings from chronic inflammation where 61 million of those cases are suffering from more than one type. Rand Corporation estimated in 2014 60% of the individual in the US had at least one chronic inflammatory condition (Pahwa & Jialal, 2018).
1.7. Effect of unregulated iNOS expression
High expression of iNOS leads to massive production of NO that results in binding to peroxynitrite that can oxidize Tetrahydrobiopterin (BH4). These significantly reduced production of physiological NO from eNOS leading to switching of activation of certain enzyme in the production of superoxide leading to major tissue damage. High level of NO also leads to hypertension by causing resistance to NO responses by the smooth muscles of the blood vessels (Chatterjee & Catravas, 2008). During diffusion of NO to the surrounding area, cells experiencing a high amount of NO will result in apoptosis and cells with a relatively low level of NO will experience inhibition of apoptosis. For this instance, if there is any tumor cell in the low level of NO will lead to high risk of developing cancer as low level of NO upregulated angiogenesis (Choudhari, Chaudhary, Bagde, Gadbail, & Joshi, 2013).
1.8. Effect of unregulated COX2
During chronic inflammation impaired COX2 causes abnormally high level of PGE2 which have commonly caused kidney dysfunction. PGE2 normally acts as an antidiuretic by controlling the vasopressin action of water absorption but at high level most nephron tubular dysfunctions, causing decrease in uptake of sodium and water resulting in development of obstructive nephropathy, kidney stones and cancer of the prostate (Nørregaard, Kwon, & Frøkiær, 2015). High level of COX2 along with PGE2 have shown to upregulate expression of metalloproteinases (MMP) and vascular endothelial growth factor (VEGF) resulting in angiogenesis leading to a higher risk of cancer if this occurs at the site of injury where chances of development of tumor cell are high (Cavaillon & Singer, 2011). High level of COX2 and PGE2 have been also correlated with multiple sclerosis because of recruitment of leukocytes to the brain leading to neuron apoptosis or axon damage (Minghetti, 2004).
1.9. Purpose of the study.
The main aim of the research is to find out if turmeric, cinnamon, and cloves have inhibitory properties against iNOS/COX2 and Nitrite levels in LPS induced inflammation in macrophages.
The objective is to carry out cell viability test using trypan blue and resazurin assay during treatment of cell using various dilution. This is to determine the maximum dilution of compounds to be used for western blotting and Griess assay without introducing cell toxicity. Griess assay is used to measure the Nitrite level. This is a more accurate level of determining the NO level as it is a very unstable molecule and has a short half-life. NO under the aerobic condition and aqueous solution its oxidize to nitrite (Ersoy et al., 2002). Higher the NO level the more likely the cells are inflamed. Cell lysis is used to extract iNOS/COX2 from the cells and BCA protein assay is used to determine how much volume of cell lysate to load in western blot to ensure equal protein concentration in each well. Finally, western blot is used to show the expression of iNOS/COX2 in various treatment of cells.
The hypothesis is when inflamed J774 cell line treated with cinnamon, cloves, and turmeric should decrease the level of expression of iNOS/COX2 and results in a significant decrease in nitrite level compared to inflamed J774 cell lines.
The null hypothesis is there is no significant difference in expression of iNOS/COX2 when inflamed J774 cell lines are treated with cinnamon, cloves, and turmeric. Nitrite levels remain the same in inflamed cell lines and inflamed cell treated with the natural compounds.
Materials
To carry out the research, the first approach is to determine the maximum dilution of the compound that can be treated to the J774 cells with high cell viability (>90%) and thus not inducing cell toxicity. The compounds (turmeric, cinnamon, and cloves) are commercially bought natural essential oil (Holland and Barrett) and dissolved in ethanol. Each well in the cell plate has a concentration of 0.80% of ethanol these were ideal for the experiment because concentration >1% showed no significant change in immunomodulatory effects on LPS induced reactive oxygen species (ROS) production (Timm, Saaby, Moesby, & Hansen, 2013).
2.1 Cell culture and Plating
Two general sets of cells were prepared, the normal cell plus the compounds and inflamed cells (induced by 1µg/ml of LPS each well) plus the compounds and were done in triplets to improve accuracy and avoid anomalous results. Positive control was no compound treated (should expect 100% viability) and negative control is the addition of Triton X (should expect 0% viability). Dilution of compounds in each well were 1:80,1:160,1:320,1:640,1:1280. The cell was cultured in Dulbecco's Modified Eagle Medium (DMEM) and when cell confluency reached above 85%, they were plated in cell culture plate or cell culture flasks.
2.2 Cell Viability Test
To determine cell viability trypan blue and resazurin assay were used. By using two different types of assay, false positive and false negative are minimized and cell viability efficiency is improved. Resazurin was diluted with DMEM to reach the concentration of 0.15mg/ml and added 20µL in each 96-cell culture plate. After incubation (5hrs) absorbance was measured at 590nm. Cell viability was calculated by OD of test sample/OD of control x 100. Trypan Blue was diluted with Phosphate buffered saline (PBS) at a ratio of 1:1 and added 20µl in each 96-cell culture plate. Cell viability was tested by taking pictures under the microscope and calculating viable cells/total cells X 100.
2.3 Cell Lysis and BCA assay.
Depending on the results of cell viability assay (when maximum dilution with >90 cell viability is obtained). The cell will be culture with one set of dilution for cell lysis and BCA assay. The cell was denatured using lysis buffer (main ingredient Tris and SDS) and Protease Inhibitor Cocktail was used to inhibit degradation of essential protein during cell lysis. Sonication is used to assist further break down of cell e.g. cell membrane.
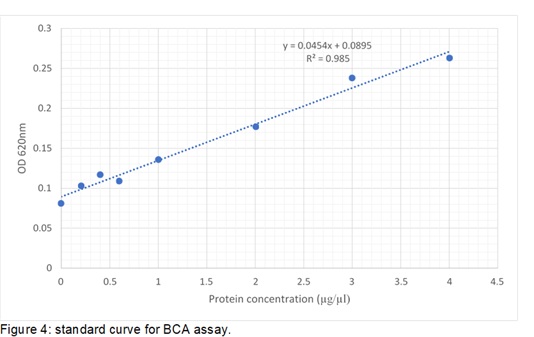
BCA assay is used to determine how much concentration of protein is available in the cell lysate. 96 well plates were used to add cell lysate and incubated with BCA reagent. The reading was taken at 620nm on the Multiskan II plate reader. Standard was made according to Figure 4, where the equation of line Y=0.0454X+0.0895 is used to calculate the protein concentration. Y presents absorbance reading and X presents protein concentration (µg/µl)
2.4. Western Blotting
Western blotting is carried out for detection of iNOS/COX2 due to its high specificity due to antibody binding, high sensitivity for protein determination and its availability in the laboratories. Each well was loaded with 20µg of cell lysate. Polyvinylidene difluoride membrane (PVDF) was used instead of Nitrocellulose during wet blotting due to its high protein binding capacity. Treatment of primary antibody were COX2 antibody produced in goat at a dilution 1:1000 and iNOS antibody produced in the mouse at a dilution 1:2500. Treatment of secondary antibody were Anti-goat IgG conjugated to Horseradish Peroxidase at a dilution 1:5000 for COX2 and Anti-mouse IgG conjugated to Horseradish Peroxidase at a dilution 1:2000 for iNOS. Enhanced Chemiluminescence (ECL) Detection was used to identify the bands and the membrane needed to be incubated for at least 60 mins.
2.5. Griess assay
Griess assay was used to measure nitrite levels (for one type of dilution depending on cell viability results) due to its availability and it was measured from the medium of cell culture plates. Absorbance was taken at 540nm on the Multiskan II plate reader. The standard curve was constructed as shown in figure 5, to determine the level of nitrite for each treatment and a bar chart was constructed. Equation of the line is Y=0.0085X+0.0303 used for calculation of nitrite levels. Y presents absorbance reading and X presents nitrite levels (µM)
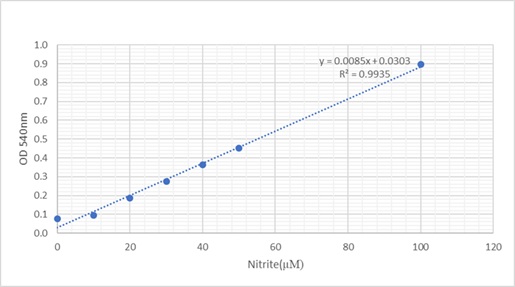
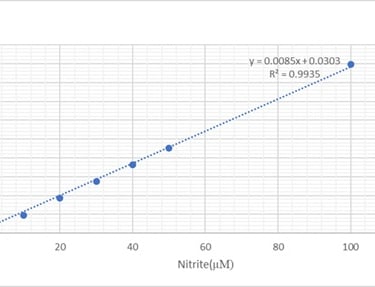
Figure 5: Standard curve for Griess assay.
Results
Overall all compounds show a decrease in expression of iNOS and COX2 these are seen by the decrease in band thickness compared to inflamed cells (control + LPS). All compounds show a decrease in Nitrite levels compared to inflamed cells. the dilution used for western blotting and Griess assay was 1/640 because this dilution both trypan blue and resazurin assay showed high viability results when tested for all compounds. This dilution is also ideal because the concentration is not so high inducing cell toxicity (explain the decrease in iNOS, COX2, and nitrite levels were due to the compounds and not because of the low viable cell) and not too low to be unable to have an effect on inflamed cells.
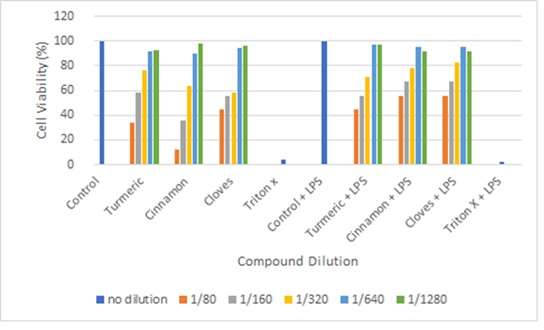
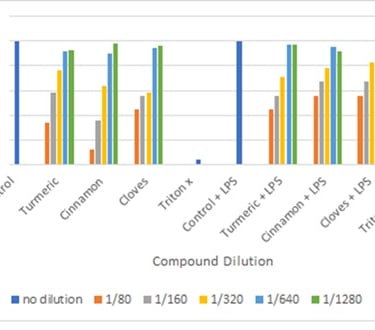
Figure 6: cell viability test for Trypan Blue. (for all compounds the general concept is as the concentration of the compound increases the cell viability decreases. Control has obtained viability of 100% while Triton X has obtained viability of >10%)
For trypan blue assay, our positive test, ie control, and control + LPS shows 100 % viability and negative test Triton X (4%) and Triton X+LPS (0%) viability. Indicating the assay was done accurate and the results produced are viable. The figure shows the average value of cell viability for each treatment done in triplets. All treatment with dilution 1/80, 1/160 and 1/320 had cell viability <90% this is not ideal to be used for western blotting or Griess assay. Both 1/640 and 1/1280 dilution have shown ≥90% viability.
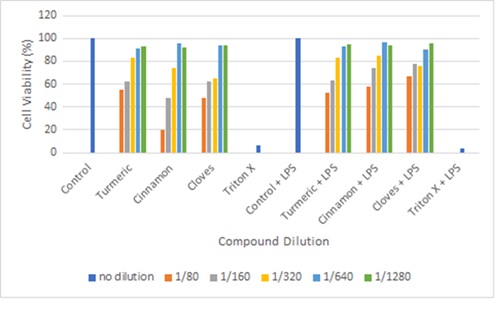
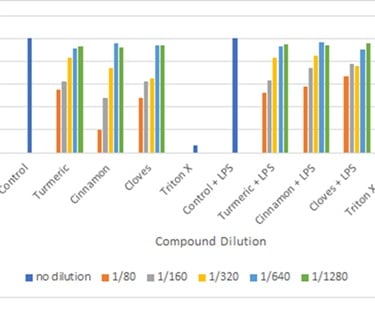
Figure 7: Cell viability test using Resazurin assay
(for resazurin assay the general concept is as the concentration of the compound increases the cell viability decreases. Control has obtained viability of 100% while Triton X has obtained viability of >10%).
When resazurin dye is oxidized, it changes color from blue to pink thus the presence of pink color indicates viable cells and blue indicates dead cells. The assay has overestimated cell viability compared to trypan blue but overall it does indicate the ideal dilution for western blot and Nitrite assay is 1/640 and 1/1280. The control and control + LPS have shown to be 100% viability while Triton X (6%) and Triton X + LPS (4%) confirming a good validity of the assay.
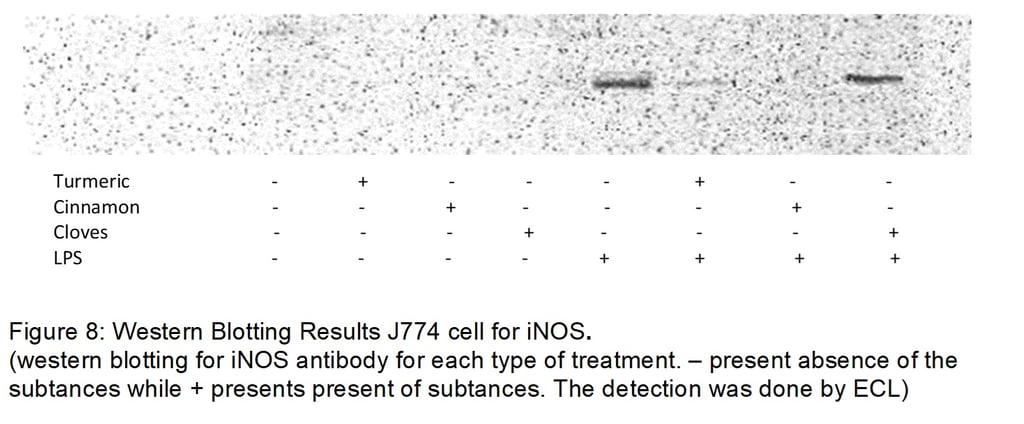
Using western blot our negative test ie controls does not show any bands. These indicate a good validity of assay for both iNOS and COX2. For control, there was no sort if treatment leading to lack of inflammation activation thus leading in no production of iNOS and COX2. Another expect is the J774 cells treated with the compounds only do not induce an inflammatory response for both assays in iNOS and COX2. Our positive test (control + LPS) present thick bands for iNOS and COX2 concluding the assay was done accurately and can also be used to compare with compound + LPS to conclude inhibitory effect (by comparing the thickness of the bands). Cinnamon shows the most inhibitory effect (no bands formation) while cloves show the least inhibitory effects. Turmeric has a better inhibitory effect than cloves and the overall result is a significant decrease in iNOS expression.
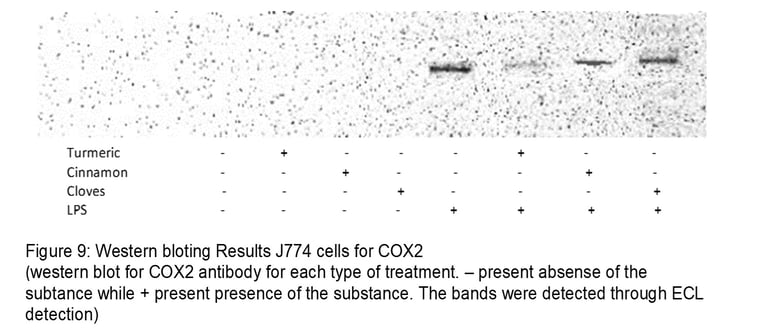
For COX2, turmeric shows the most inhibitory effect while cloves show the least inhibitory effect. As there is very little band formation for turmeric. Cinnamon also has to some extent an inhibitory effect on COX2. Overall all turmeric seems to be the most efficient in controlling inflammation as it has significantly inhibited both iNOS and COX2. Cinnamon seems to effect iNOS better than COX2. Cloves have the least effect on inflammation as it shows a slight decrease in the bands compared to control +LPS for both iNOS and COX2.
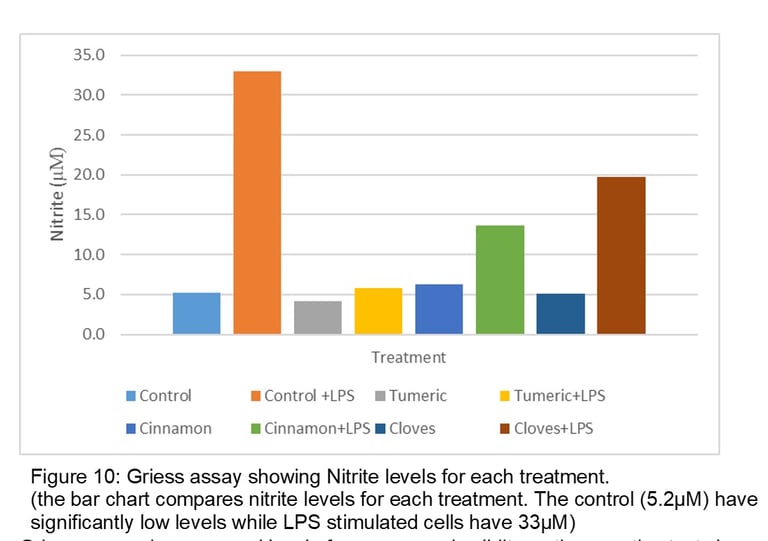
Griess assay shows a good level of accuracy and validity as the negative test shows a low level of Nitrite. Low expression of iNOS leads to low production of NO resulting in low Nitrite levels and vice versa. The positive test (control +LPS) has the most nitrite levels indicating success in inducing inflammation. It is also used to compare the decrease of nitrite when treating inflamed cells with compounds. The results also show a low level of Nitrite in J774 cells treated with only turmeric (4.2µM), Cinnamon (6.3µM) and Cloves (5.1µM) which shows similarity to figure 8. Turmeric has the most effect (with a difference of 27.2µM when compared with Control +LPS) and cloves have the least effect (with a difference of 13.3 µM when compared with Control +LPS on Nitrite levels). One key finding is despite cinnamon have no bands thus completely inhibits iNOS expression this does not lead low levels of nitrite (difference of 19.4µM when compared with control +LPS).
Discussion
Overall the results confirm the compounds do have an inhibitory effect of iNOS, COX2 and Nitrite levels in LPS induced inflammation in J774 cell lines thus may have clinical importance for further treatment. The Hypothesis is accepted and the null hypothesis is rejected.
LPS was the ideal treatment as it induces myd88 dependent pathway of inflammation signaling. Macrophages were chosen as they are the first type of leukocyte found at the site of inflammation and has a major part in initiation, maintenance, and termination of inflammation. During these processes, they also play a role in innate and adaptive immune response (Fujiwara & Kobayashi, 2005).
Ethanol was chosen as it displays the least cell toxicity and produced most viable cell compared to, dimethyl sulfoxide (DMSO) and dimethylformamide (DMF) in RAW-264.7 cell lines (Jamalzadeh et al., 2016).
Using Figure 8, turmeric (the active ingredient is curcumin) may affect the gene expression of iNOS by inhibiting its mRNA and degrading the iNOS protein itself resulting is very thin band formation (Jung et al., 2006). In macrophages proteasome is inhibited by lactacystin and ubiquitination is inhibited by Benzyloxycarbonylleucylleucylleucinal (MG123) these greatly increases the amount of iNOS. It has been suggested by inactivating of MG123 and Lactacystin either by dephosphorylation or enhancing the degradation of the molecules leading to high expression of ubiquitination and proteolysis of iNOS resulting in a low level of iNOS and eventually NO and Nitrite levels (Ben et al., 2011). Curcumin terminates the degradation of IkB complex by inhibiting its proteasome and prevents the heterodimerization of IKK complex both preventing the cytosolic release of p50/p65-RelA complex leading to a decrease in expression of iNOS/COX2. Curcumin has shown to be able to prevent the cytosolic release of P65 preventing formation of p65-Rel A complex (Pan, Lin-Shiau, & Lin). Curcumin can inhibit the expression of mRNA and translation of COX2 resulting in decrease in protein expression resulting in thinner bands. The mechanism of inhibition is not well implied (Goel, Boland, & Chauhan, 2001).
Cinnamon most anti-inflammatory active ingredient cinnamaldehyde has shown to downregulate IkB degradation by dephosphorylation and NF-kb protein (protein complex in the NF-kb pathway that enhances transcription of NF-Kb genes) by enhancing ubiquitination. By effecting NF-kb protein all 3 pathways of NF-kb are downregulated these prevents transcription of INOS/COX2 resulting in no bands of iNOS (Liao et al.). The unexpected nitrite level in cinnamon despite no translation of iNOS is due to its ability to produce NO via enhancement of expression of eNOS pathway (Raffai et al., 2014). One suggestion is it enhances Protein Kinase B (AKT) signalling pathway leading to transcription of AKT, that have a key role in the phosphorylation of eNOS at serine 1177 to generate NO (Lu et al., 2018). Cinnamaldehyde also increases the production of nNOS by upregulating expression Transient receptor potential ankyrin-1 (TRPA1) leading to gene activation of nNOS (Aubdool et al., 2016). The reason why no bands formation for iNOS while thin bands formation of COX2 despite its high activation of inhibition in NF-kb pathway is the cinnamon ability to upregulated transforming growth factor beta (TGF-β) (Jana et al., 2013). TGF-β enhances instability of iNOS mRNA by upregulating the binding of TCF11/MafG to the promoter region leading to degradation of mRNA and eventually inhibition in synthesis of iNOS (Berg et al., 2007). TGF-β has also been shown to upregulate COX2 by expression of Smad (Fang, Chang, Cheng, Leung, & Sun, 2014).
Eugenol is the main active ingredient in cloves and it has been shown to have an inhibitory effect on iNOS expression when stimulated with LPS but no effect on iNOS protein without LPS stimulation and the reason is unknown. The mechanism of how eugenol downregulated is not very well known as it has proven that it has no effect of IkB protein. Its anti-inflammatory effect is by inhibiting the formation of peroxynitrite (Li et al., 2006). Studies have shown Eugenol downregulates the NF-kb pathway by decreasing the DNA affinity to the NF-kb protein and expressing low levels of P65 resulting in a decrease in iNOS (decrease in nitrite level) and COX2 but as not much compared to turmeric and cinnamon (Yeh et al., 2011). Another reason for the low inhibitory activity of iNOS/COX2 compared to cinnamon and turmeric is a higher concentration may be required to have any significant inhibitory effect.
All Compounds have known to inhibit activity Activator protein 1 (AP-1), which is a transcription factor that regulates genes expression of various cytokines including iNOS and COX2 (Fujioka et al., 2004).
Importance of the results is awareness of the compounds ability to regulate inflammation by downregulation of iNOS and COX2. Thus, the compounds downregulate iNOS and COX2 have high ability in the prevention of insulin resistance. Studies have shown a high correlation between upregulation of iNOS (Fujimoto et al., 2005) and COX2 causes decrease in insulin sensitivity in type 2 diabetes and obesity-related insulin resistance (Persaud, Burns, Belin, & Jones, 2004). The mechanism is not fully understood but one suggestion is the compounds enhance insulin sensitivity molecules such as Thiazolidinediones (that also have an inhibitory effect of iNOS) (Dang et al., 2013). Heart disease such as atherosclerosis has been shown to upregulated iNOS and loss ability in physiological production of NO through eNOS. Compounds ideally cinnamon may have the ability to inhibit the progression of atherosclerosis and aid in the prevention of the disease (Kawashima & Yokoyama, 2004). It may also help us to understand if the compounds ability to inhibit NO by iNOS production during hypertrophy that has been shown to contribute to myocardial dysfunction (Förstermann & Sessa, 2012). The compounds may be used for anti-cancer activities as it is well known most cancer express attributes of inflammation. Certain cancer eg colorectal cancer has been shown to express and induced by the high amount of COX2 (Ferrandez, Prescott, & Burt, 2003) and while Triple-Negative Breast Cancer express a high level of iNOS (Jin et al., 2015).
Many studies have found out the compounds have inhibitory effects for iNOS and COX2 for a wide variety of cells. Curcumin has inhibited iNOS in Microglial cells induced by Lipoteichoic acid. that play a key role in chronic neuroinflammation that induces diseases such as Parkinson’s disease, Huntington’s disease, stroke and multiple sclerosis (Yu et al., 2018). It also degrades iNOS/COX2 in RAW 264.7 cells (Ben et al., 2011), CD-5 positive B-Cells in chronic lymphocytic leukaemia (Shanbhag, 2017), HT-29 human colon cancer cells (Goel et al., 2001) and various gastrointestinal cell lines such as SK-GT-4, SCC450, IEC-18 and HCA-7 (Zhang, Altorki, Mestre, Subbaramaiah, & Dannenberg, 1999). Cinnamon aldehyde has also been shown to inhibit iNOS/COX2 level in RAW 264.7(Lu et al., 2018), human peripheral blood mononuclear cells, THP1XBlue human monocytic cell line (Roth-Walter et al., 2014), BV-2 microglial cells (Y.-F. Chen et al., 2016). Eugenol has also shown to inhibit iNOS and COX2 in RAW264.7 cells, human dental pulp fibroblasts and HeLa cells (Barboza, da Silva Maia Bezerra Filho, Silva, Medeiros, & de Sousa, 2018). No research was found that the compounds did not affect iNOS and COX2 levels.
One of the common diseases that arise from acute and chronic inflammation in rheumatoid arthritis with no cure. Current treatments such as Nonsteroidal anti-inflammatory drugs (NSAIDs), Disease-modifying antirheumatic drugs (DMARDs) and corticosteroids are costly and have an adverse side effect. The compounds may have the ability to cure the disease and may able to provide a better alternative with less health risk and side effect with better cost (Bortoluzzi et al., 2018). Curcumin has been proven significant decrease visual analog scale, Western Ontario and McMaster Universities Osteoarthritis Index (WOMAC) scores and Lequesne’s pain functional index (LPFI) score in 40 human subjects suffering from arthritis in a randomized double-blind placebo-controlled trial (Hewlings & Kalman, 2017). A study done on randomized 30 human subject both cinnamaldehyde and eugenol have shown to significantly reduce the iNOS level and reduce the effect of oxidative stress due to excess NO and assisted in the management of the disease in arthritis (Mateen et al., 2019). Another disease with minimum treatment and no cure caused by chronic neuroinflammation is Alzheimer disease that has been known expressed high levels of iNOS and COX2. Various research has confirmed curcumin significantly reduces the risk of development of Alzheimer disease. For eg India (due to their high use of turmeric in their diets) are 4.4 less likely than United Stated in developing Alzheimer disease in individuals ages 70-70yrs (Mishra & Palanivelu, 2008). By inhibition of oxidative stress eg inhibiting the formation of peroxynitrite by eugenol can prevent neuronal death in neuronal degenerative disease such as Parkinson disease (Kim, Kim, Rhie, & Yoon, 2015). Compounds may have potential use for treatment for bowel inflammatory diseases such as Crohn disease where it has no cure, asthma, type 2 diabetes, and certain cancers but further clinical research has to be done for concrete evidence.
Since the study was a vitro experiment, it lacks the basic understanding of xenobiotic metabolism, molecular interaction/ toxicity between different types of cells, tissue or organ system in the body, consequence of long term exposure that impact the effectiveness of the treatment (Aslantürk, 2017). Trypan blue measure integrity of viable cells and factors such as poor dispersion of cell and not using hemacytometer can introduce error. Resazurin assay measures the cell metabolism and error such as interferences from the compounds can occur (Aslantürk, 2017). For Griess assay excessive NADPH produce in the cell may affect the sensitivity of the assay because nitrite reductance is enhanced by NADPH. The media was also not deproteinized that may interfere with absorbance reading (Csonka et al., 2015).
Further improvement of the study is using western blot/Griess assay with a wide range of dilution of the compounds in the inflamed cells. As the concentration (with high cell viability) increases the expression of iNOS and COX2 should decrease. Western blot of NF-kb, AP-1, and IkB protein can be carried out to confirm the effect of the compounds iNOS/COX2 transcription pathway. During Western blot, B-actin is used to ensure equal protein is loaded across the cell. For analysing the bands, densitometry analysis is ideal as it enables to accurately compare the thickness of the band without background interferences. Cell counting is also idea before cell plating to ensure an equal number of cells in each well. A separate protocol to determine ethanol toxicity is recommended to ensure accurate preparation of the compounds. More runs are required to improve the accuracy of the results.
Further research involves working with other proinflammatory cytokines such as TGF-α that have a major role in pathological pain in chronic inflammation, anti-inflammatory cytokines such as interleukin 4 (IL-4) or interleukin (IL-10)(J.-M. Zhang & An, 2007). Analysis of other types of immune cells such as dendritic, neutrophil and T lymphocytes. Research can be done with inflamed cells induced by DAMPS, Poly IC (TRIF dependent) or lipoteichoic acid (derived from Gram-positive bacteria unlike LPS that derived from Gram-negative).
In Conclusion, the compounds have the ability to inhibited iNOS/COX2 but further Vitro/vivo research is required due to the complexity of inflammation before entering clinical trials for treatment.
References
1. Aslantürk, Ö. S. (2017). In Vitro Cytotoxicity and Cell Viability Assays: Principles, Advantages, and Disadvantages. In: IntechOpen.
2. Aubdool, A. A., Kodji, X., Abdul-Kader, N., Heads, R., Fernandes, E. S., Bevan, S., & Brain, S. D. (2016). TRPA1 activation leads to neurogenic vasodilatation: involvement of reactive oxygen nitrogen species in addition to CGRP and NO. Br J Pharmacol, 173(15), 2419-2433.
3. Barboza, J. N., da Silva Maia Bezerra Filho, C., Silva, R. O., Medeiros, J. V. R., & de Sousa, D. P. (2018). An Overview on the Anti-inflammatory Potential and Antioxidant Profile of Eugenol. Oxidative medicine and cellular longevity, 2018, 3957262-3957262.
4. Ben, P., Liu, J., Lu, C., Xu, Y., Xin, Y., Fu, J., . . . Yin, Z. (2011). Curcumin promotes degradation of inducible nitric oxide synthase and suppresses its enzyme activity in RAW 264.7 cells. International Immunopharmacology, 11(2), 179-186.
5. Berg, D. T., Gupta, A., Richardson, M. A., O'Brien, L. A., Calnek, D., & Grinnell, B. W. (2007). Negative regulation of inducible nitric-oxide synthase expression mediated through transforming growth factor-beta-dependent modulation of transcription factor TCF11. J Biol Chem, 282(51), 36837-36844.
6. Bortoluzzi, A., Furini, F., Generali, E., Silvagni, E., Luciano, N., & Scire, C. A. (2018). One year in review 2018: novelties in the treatment of rheumatoid arthritis. Clin Exp Rheumatol, 36(3), 347-361.
7. Cavaillon, J.-M. e., & Singer. (2011). M. e. Inflammation : From Molecular and Cellular Mechanisms to the Clinic (1st ed.).
8. Chatterjee, A., & Catravas, J. D. (2008). Endothelial nitric oxide (NO) and its pathophysiologic regulation. Vascular Pharmacology, 49(4), 134-140.
9. Chen, L., Deng, H., Cui, H., Fang, J., Zuo, Z., Deng, J., . . . Zhao, L. (2017). Inflammatory responses and inflammation-associated diseases in organs. Oncotarget, 9(6), 7204-7218.
10. Chen, Y.-F., Wang, Y.-W., Huang, W.-S., Lee, M.-M., Gibson Wood, W., Leung, Y.-M., & Tsai, H.-Y. (2016). Trans-Cinnamaldehyde, An Essential Oil in Cinnamon Powder, Ameliorates Cerebral Ischemia-Induced Brain Injury via Inhibition of Neuroinflammation Through Attenuation of iNOS, COX-2 Expression and NFκ-B Signaling Pathway (Vol. 18).
11. Choudhari, S. K., Chaudhary, M., Bagde, S., Gadbail, A. R., & Joshi, V. (2013). Nitric oxide and cancer: a review. World journal of surgical oncology, 11, 118-118. doi:10.1186/1477-7819-11-118
12. Csonka, C., Páli, T., Bencsik, P., Görbe, A., Ferdinandy, P., & Csont, T. (2015). Measurement of NO in biological samples. British journal of pharmacology, 172(6), 1620-1632.
13. Dang, H. Z., Li, R. S., Wang, H., Ren, Y., Sun, H. Y., Yang, J. D., & Wang, P. W. (2013). [Effects of curcumin on expression of PI3K and p-pI3K in hippocampus of AD mice]. Zhongguo Zhong Yao Za Zhi, 38(9), 1295-1299.
14. Du Clos, T. W. (2000). Function of C-reactive protein. Ann Med, 32(4), 274-278.
15. Ersoy, Y., Özerol, E., Baysal, Ö., Temel, I., MacWalter, R. S., Meral, Ü., & Altay, Z. E. (2002). Serum nitrate and nitrite levels in patients with rheumatoid arthritis, ankylosing spondylitis, and osteoarthritis. Annals of the Rheumatic Diseases, 61(1), 76.
16. Fang, L., Chang, H. M., Cheng, J. C., Leung, P. C., & Sun, Y. P. (2014). TGF-beta1 induces COX-2 expression and PGE2 production in human granulosa cells through Smad signaling pathways. J Clin Endocrinol Metab, 99(7), E1217-1226.
17. Ferrandez, A., Prescott, S., & Burt, R. W. (2003). COX-2 and colorectal cancer. Curr Pharm Des, 9(27), 2229-2251.
18. Foster, J. R. (2001). The functions of cytokines and their uses in toxicology. International journal of experimental pathology, 82(3), 171-192.
19. Fujimoto, M., Shimizu, N., Kunii, K., Martyn, J. A., Ueki, K., & Kaneki, M. (2005). A role for iNOS in fasting hyperglycemia and impaired insulin signaling in the liver of obese diabetic mice. Diabetes, 54(5), 1340-1348.
20. Fujioka, S., Niu, J., Schmidt, C., Sclabas, G. M., Peng, B., Uwagawa, T., . . . Chiao, P. J. (2004). NF-κB and AP-1 Connection: Mechanism of NF-κB-Dependent Regulation of AP-1 Activity. Molecular and Cellular Biology, 24(17), 7806.
21. Fujiwara, N., & Kobayashi, K. (2005). Macrophages in inflammation. Curr Drug Targets Inflamm Allergy, 4(3), 281-286.
22. Förstermann, U., & Sessa, W. C. (2012). Nitric oxide synthases: regulation and function. European heart journal, 33(7), 829-837d.
23. Gilmore, T. D. (2006). Introduction to NF-kappaB: players, pathways, perspectives. Oncogene, 25(51), 6680-6684.
24. Goel, A., Boland, C. R., & Chauhan, D. P. (2001). Specific inhibition of cyclooxygenase-2 (COX-2) expression by dietary curcumin in HT-29 human colon cancer cells. Cancer Letters, 172(2), 111-118.
25. Hewlings, S. J., & Kalman, D. S. (2017). Curcumin: A Review of Its' Effects on Human Health. Foods (Basel, Switzerland), 6(10), 92.
26. Hong, J., Bose, M., Ju, J., Ryu, J. H., Chen, X., Sang, S., . . . Yang, C. S. (2004). Modulation of arachidonic acid metabolism by curcumin and related beta-diketone derivatives: effects on cytosolic phospholipase A(2), cyclooxygenases and 5-lipoxygenase. Carcinogenesis, 25(9), 1671-1679.
27. Hun Kim, B., Lee, Y. G., Lee, J., Young Lee, J., & Cho, J. (2010). Regulatory Effect of Cinnamaldehyde on Monocyte/Macrophage-Mediated Inflammatory Responses (Vol. 2010).
28. Jamalzadeh, L., Ghafoori, H., Sariri, R., Rabuti, H., Nasirzade, J., Hasani, H., & Reza Aghamaali, M. (2016). Cytotoxic Effects of Some Common Organic Solvents on MCF-7, RAW-264.7 and Human Umbilical Vein Endothelial Cells (Vol. In press).
29. Jana, A., Modi, K. K., Roy, A., Anderson, J. A., van Breemen, R. B., & Pahan, K. (2013). Up-regulation of neurotrophic factors by cinnamon and its metabolite sodium benzoate: therapeutic implications for neurodegenerative disorders. J Neuroimmune Pharmacol, 8(3), 739-755.
30. Jin, Z., Wang, W., Jiang, N., Zhang, L., Li, Y., Xu, X., . . . Chen, B. (2015). Clinical Implications of iNOS Levels in Triple-Negative Breast Cancer Responding to Neoadjuvant Chemotherapy. PLoS One, 10(7), e0130286.
31. Jung, K. K., Lee, H. S., Cho, J. Y., Shin, W. C., Rhee, M. H., Kim, T. G., . . . Kang, S. Y. (2006). Inhibitory effect of curcumin on nitric oxide production from lipopolysaccharide-activated primary microglia. Life Sciences, 79(21), 2022-2031.
32. Kawashima, S., & Yokoyama, M. (2004). Dysfunction of endothelial nitric oxide synthase and atherosclerosis. Arterioscler Thromb Vasc Biol, 24(6), 998-1005.
33. Kim, G. H., Kim, J. E., Rhie, S. J., & Yoon, S. (2015). The Role of Oxidative Stress in Neurodegenerative Diseases. Experimental neurobiology, 24(4), 325-340.
34. Li, W., Tsubouchi, R., Qiao, S., Haneda, M., Murakami, K., & Yoshino, M. (2006). Inhibitory action of eugenol compounds on the production of nitric oxide in RAW264.7 macrophages. Biomed Res, 27(2), 69-74.
35. Liao, J.-C., Deng, J.-S., Chiu, C.-S., Hou, W.-C., Huang, S.-S., Shie, P.-H., & Huang, G.-J.
36. Lu, Z., Lili, W., Ximing, L., Hisae, Y., Katsumi, I., Hiroko, N., . . . Ming, G. (2018). Trans-cinnamaldehyde promotes nitric oxide release via the protein kinase-B/v-Akt murine thymoma viral oncogene -endothelial nitric oxide synthase pathway to alleviate hypertension in SHR. Cg-Leprcp/NDmcr rats. Journal of Traditional Chinese Medicine, 38(4), 548-555.
37. Mateen, S., Rehman, M. T., Shahzad, S., Naeem, S. S., Faizy, A. F., Khan, A. Q., . . . Moin, S. (2019). Anti-oxidant and anti-inflammatory effects of cinnamaldehyde and eugenol on mononuclear cells of rheumatoid arthritis patients. European Journal of Pharmacology, 852, 14-24.
38. Minghetti, L. (2004). Cyclooxygenase-2 (COX-2) in Inflammatory and Degenerative Brain Diseases. Journal of Neuropathology & Experimental Neurology, 63(9), 901-910.
39. Mishra, S., & Palanivelu, K. (2008). The effect of curcumin (turmeric) on Alzheimer's disease: An overview. Annals of Indian Academy of Neurology, 11(1), 13-19.
40. Mogensen, T. H. (2009). Pathogen recognition and inflammatory signaling in innate immune defenses. Clinical microbiology reviews, 22(2), 240-273.
41. Nakanishi, M., & Rosenberg, D. W. (2013). Multifaceted roles of PGE2 in inflammation and cancer. Seminars in immunopathology, 35(2), 123-137.
42. Nørregaard, R., Kwon, T.-H., & Frøkiær, J. (2015). Physiology and pathophysiology of cyclooxygenase-2 and prostaglandin E2 in the kidney. Kidney research and clinical practice, 34(4), 194-200.
43. Pahwa, R., & Jialal, I. (2018). Chronic Inflammation. In. Treasure Island: StatPearls Publishing.
44. Pan, M.-H., Lin-Shiau, S.-Y., & Lin, J.-K.
45. Persaud, S. J., Burns, C. J., Belin, V. D., & Jones, P. M. (2004). Glucose-Induced Regulation of COX-2 Expression in Human Islets of Langerhans. Diabetes, 53(suppl 1), S190.
46. Raffai, G., Kim, B., Park, S., Khang, G., Lee, D., & Vanhoutte, P. M. (2014). Cinnamaldehyde and cinnamaldehyde-containing micelles induce relaxation of isolated porcine coronary arteries: role of nitric oxide and calcium. International journal of nanomedicine, 9, 2557-2566.
47. Ricciotti, E., & FitzGerald, G. A. (2011). Prostaglandins and inflammation. Arteriosclerosis, thrombosis, and vascular biology, 31(5), 986-1000.
48. Roth-Walter, F., Moskovskich, A., Gomez-Casado, C., Diaz-Perales, A., Oida, K., Singer, J., . . . Jensen-Jarolim, E. (2014). Immune suppressive effect of cinnamaldehyde due to inhibition of proliferation and induction of apoptosis in immune cells: implications in cancer. PloS one, 9(10), e108402-e108402.
49. Shanbhag, V. K. L. (2017). Curcumin in chronic lymphocytic leukemia – A review. Asian Pacific Journal of Tropical Biomedicine, 7(6), 505-512.
50. Shu, Q., Chen, L., He, W.-j., Zhou, H.-j., & Liang, F.-x. (2016). A review of inflammatory signaling pathway regulated by acupuncture. World Journal of Acupuncture - Moxibustion, 26(2), 63-69.
51. Straub, R. H., & Schradin, C. (2016). Chronic inflammatory systemic diseases: An evolutionary trade-off between acutely beneficial but chronically harmful programs. Evolution, medicine, and public health, 2016(1), 37-51.
52. Timm, M., Saaby, L., Moesby, L., & Hansen, E. W. (2013). Considerations regarding use of solvents in in vitro cell based assays. Cytotechnology, 65(5), 887-894.
53. Wallace, J. L. (2005). Nitric oxide as a regulator of inflammatory processes. Memórias do Instituto Oswaldo Cruz, 100, 5-9.
54. Wassell, J. (2000). Haptoglobin: function and polymorphism. Clin Lab, 46(11-12), 547-552.
55. Yeh, J. L., Hsu, J. H., Hong, Y. S., Wu, J. R., Liang, J. C., Wu, B. N., . . . Liou, S. F. (2011). Eugenolol and glyceryl-isoeugenol suppress LPS-induced iNOS expression by down-regulating NF-kappaB AND AP-1 through inhibition of MAPKS and AKT/IkappaBalpha signaling pathways in macrophages. Int J Immunopathol Pharmacol, 24(2), 345-356.
56. Yu, Y., Shen, Q., Lai, Y., Park, S. Y., Ou, X., Lin, D., . . . Zhang, W. (2018). Anti-inflammatory Effects of Curcumin in Microglial Cells. Frontiers in Pharmacology, 9(386).
57. Zhang, F., Altorki, N. K., Mestre, J. R., Subbaramaiah, K., & Dannenberg, A. J. (1999). Curcumin inhibits cyclooxygenase-2 transcription in bile acid- and phorbol ester-treated human gastrointestinal epithelial cells. Carcinogenesis, 20(3), 445-451.
58. Zhang, J.-M., & An, J. (2007). Cytokines, inflammation, and pain. International anesthesiology clinics, 45(2), 27-37.
